転移RNA

リボソームは、一連の伝令RNA(en: Messenger RNA)を読み取り、転移RNA(en: Transfer RNA (TRNA))に結びついたアミノ酸から所定のタンパク質を組み立てる。
小胞体に分泌されるタンパク質を組み立てているリボソーム
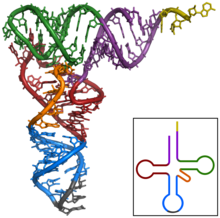
転移RNAの分子モデル

転移RNAの構造(クローバーリーフ)
転移RNA(てんい-、transfer RNA)は73〜93塩基の長さの小さなRNAである。リボソームのタンパク質合成部位でmRNA上の塩基配列(コドン)を認識し、対応するアミノ酸を合成中のポリペプチド鎖に転移させるためのアダプター分子である。運搬RNA、トランスファーRNAなどとも呼ぶが、通常tRNAと略記される。
目次
1 構造
2 アンチコドン
3 アミノアシル化
4 tRNA遺伝子
5 訳語について
6 関連項目
7 参考文献
8 外部リンク
構造
通常は、D・アンチコドン・Tという3つのアームを持つクローバーリーフと呼ばれる二次構造を持ち、これが折り畳まれて3次元的にはL字型になる。L字の長い側の先端にはアンチコドンがありmRNA上のコドンと対合する。短い側の先端にはアミノ酸が結合しポリペプチド合成に用いられる。tRNAの塩基は化学修飾を受けているものも多く、なかでもメチル化は頻繁に見られる。
- アクセプターステム
- L字型の短い側に相当する。一次構造上の両末端が対合しているが、ゆらぎ塩基対を含む場合がある。5'末端はリン酸基を持つ。3'末端側はCCAの3塩基が突出し、末端のアデノシン残基にアミノ酸が共有結合する。CCA配列は殆どの真正細菌ではtRNA本体と同様に遺伝子から転写されるが、真核生物と古細菌においては転写後にCCA付加酵素によって付加される。古細菌ではクラスI-CCA付加酵素、真核生物(と一部の真正細菌)ではクラスII-CCA付加酵素によって行われる。
- Dアーム
- L字型の長い側の基部に相当し、アンチコドンアームに対して上流側のステムループである。Dループ-Tループの相互作用は三次構造形成に重要である。アミノアシルtRNA合成酵素によって認識される部位だと考えられている[要出典]。修飾塩基としてジヒドロウリジン(D)を含むことが多い。
- アンチコドンアーム
- L字型の長い側の先端に相当するステムループであり、ループ中にコドンと対合するアンチコドンが存在する。アンチコドンの1文字目には様々な修飾塩基が見られ、コドン認識に重要な役割を担っている。アンチコドンの3'側に隣接する37位も頻繁に修飾を受ける。
- Tアーム
- L字型の関節部に相当し、アンチコドンアームに対して下流側のステムループである。リボソームによって認識される部位だと考えられている[要出典]。典型的なtRNAではループ中にTΨCという保存配列があり、RNAであるにも関わらず修飾塩基としてチミジン(T, 正確にはリボチミジン(rT)または5-メチルウリジン(m5U))を含むことが多い。T、Ψ(シュードウリジン)とも生物種によってはそれらの類縁体になっていることもある。
セレノシステイン-tRNAと、ピロリシン-tRNA は例外的に他のtRNAにない様々な特徴を持つ。
アンチコドン
この節の正確性に疑問が呈されています。問題箇所に信頼できる情報源を示して、記事の改善にご協力ください。議論はノートを参照してください。(2008年3月) |
mRNA上のコドンと対合する3塩基をアンチコドンと呼ぶ。例えばAAAというコドンはリジンをコードしているが、これに対応するリジンtRNAのアンチコドンはUUUとなっている。しかしこの対応関係は1対1とは限らず、1つのアンチコドンが同じアミノ酸をコードする複数のコドンを認識する場合がある。仮に1対1だとすれば61種のtRNAが必要になるが、通常はこれよりも少ない種類のtRNAしか存在しない。
アンチコドンの1文字目は化学修飾によりイノシン(I)またはシュードウリジン(Ψ)になっている場合がある。これらの修飾塩基は複数の塩基と水素結合を形成できるため、こうしたtRNAは3文字目だけが異なる複数のコドンを認識できる。例えばグリシンをコードするコドンは、GGU・GGC・GGA・GGGの4つだが、アンチコドンの1文字目が化学修飾されていれば1つのtRNAで4つのコドンを読むことが可能である。
tRNAはそれぞれ特定のアミノ酸としか結合しないが、遺伝暗号が縮重しているため、異なるアンチコドンを持つtRNAが同じアミノ酸と結合する場合がある。1つのアミノ酸に対して2種類以上のtRNAが存在し、1つのtRNAは複数のコドンに対応しうるため、30から40種のtRNAが1つの翻訳系で使われる。
アミノアシル化
tRNAの3'末端にあるCCAのアデノシン残基には、tRNAごとに特定のアミノ酸が結合してアミノアシルtRNAとなる。この反応をアミノアシル化といい、アミノアシルtRNA合成酵素によって触媒される。通常はアミノ酸ごとに1種類のアミノアシル化tRNA合成酵素が存在しており、アンチコドンが異なる複数のtRNAを1種の酵素が認識してアミノアシル化を触媒している。コドンとアミノ酸の正確な対応には、tRNAとアミノアシルtRNA合成酵素の特異的な相互作用が必須となる。この対応関係はアンチコドンだけを認識して決定しているわけではないらしい。
tRNA遺伝子
ゲノム中のtRNA遺伝子の数は生物により様々である。線虫C. elegansの核ゲノムには全部で19000遺伝子があるが、そのうち659がtRNAをコードしている[1]。出芽酵母では275である。ヒトでは497個が知られており、アンチコドンごとに整理すると49種となる。またヒトのゲノム中にはtRNA由来の偽遺伝子が324個見つかっている[2]。
真核生物では、mRNAがRNAポリメラーゼIIにより転写されるのに対し、tRNAはRNAポリメラーゼIIIによって転写される。その後Pre-tRNA スプライシングや塩基修飾を経て成熟型のtRNA分子へと加工される。
訳語について
学術用語集では植物学編・遺伝学編が「転移RNA(運搬RNA)」、動物学編が「運搬RNA」としている(すべて増訂版)。JISの生体工学用語(K3610)では「転移RNA」である。一般には「転移RNA」の方が好んで用いられる傾向にあるが、高校教育では「運搬RNA」が用いられている。
関連項目
- 修飾塩基
- ゆらぎ説
参考文献
^ Hartwell LH, Hood L, Goldberg ML, Reynolds AE, Silver LM, Veres RC. (2004). Genetics: From Genes to Genomes 2nd ed. McGraw-Hill: New York, NY. p 264.
^ Lander E. et al. (2001年). “Initial sequencing and analysis of the human genome”. Nature 409 (6822): 860-921. PMID 11237011.
外部リンク
- tRNAdb 2009: compilation of tRNA sequences and tRNA genes
- Compilation of tRNA sequences and sequences of tRNA genes
- Genomic tRNA database
- Compilation of mammalian mitochondrial tRNA genes
- tRNADB-CE: tRNA gene database curated manually by experts
| ||||||||||||||


