進化

生物は共通祖先から進化し、多様化してきた。
進化(しんか、羅: evolutio、英: evolution)は、生物の形質が世代を経る中で変化していく現象のことである[1][2]。
目次
1 定義
2 進化の証拠
2.1 古生物学
2.1.1 ミッシング・リンク
2.2 生物地理学から
2.3 比較解剖学から
2.3.1 相似と相同
2.3.2 痕跡
2.3.3 不合理な形態
2.4 系統分類学から
2.5 発生生物学から
2.6 観察された進化
2.6.1 実験進化
3 進化のしくみ
3.1 遺伝的変異
3.2 遺伝子の頻度変化
3.2.1 自然選択
3.2.2 遺伝的浮動
4 進化の速度
4.1 形態の進化
4.2 分子進化
5 大進化
5.1 種分化
5.1.1 断続平衡説
6 進化に関する誤解
7 生物学以外での「進化」概念
8 脚注
9 参考文献
10 関連項目
11 外部リンク
定義

眼の進化
進化とは、生物個体群の性質が、世代を経るにつれて変化する現象である[2][1]。また、その背景にある遺伝的変化を重視し、個体群内の遺伝子頻度の変化として定義されることもある[3][4]。この定義により、成長や変態のような個体の発生上の変化は進化に含まれない[1][2]。
また狭義に、種以上のレベルでの変化のみを進化とみなすこともあるが、一般的ではない[3]。逆に、文化的伝達による累積的変化や生物群集の変化をも広く進化と呼ぶこともある[3]。日常表現としては単なる「変化」の同義語として使われることも多く、恒星や政治体制が「進化」するということもあるが、これは生物学でいう進化とは異なる[4]。
進化過程である器官が単純化したり、縮小したりすることを退化というが[3]、これもあくまで進化の一つである。退化は進化の対義語ではない。
進化の証拠
生物は不変のものではなく、共通祖先から長大な年月の間に次第に変化して現生の複雑で多様な生物が生じたということが、膨大な証拠から分かっている[5][6]。
進化論は、チャールズ・ダーウィンなど複数の博物学者が動物や植物の分類学的な洞察から導きだした仮説から始まった。現在の自然科学ではこの説を裏付ける証拠が、形態学、遺伝学、比較発生学、分子生物学などさまざまな分野から提出されており、進化はほぼ確実に起こってきた事実である、と生物学者・科学者からは認められている[5][6][7]。
古生物学
進化をはっきりと示す化石証拠はダーウィンの時代には乏しかったが、現在では豊富に存在する。まず全体的なパターンとして、単純で祖先的と思われる生物は古い地層からも見つかるが、複雑で現生種に似た生物は新しい地層からしか見つからない[8]。
化石証拠の豊富な生物については、化石を年代順に並べることで、特定の系統の進化を復元することもできる。プランクトンは死骸が古いものから順に連続的に堆積していくので、このような研究が容易であり、有孔虫や放散虫、珪藻の形態が徐々に進化し、時には種分化する過程が確認できる[9][10]。プランクトン以外にも、三葉虫の尾節の数の進化を示す一連の化石などがある[10]。
ミッシング・リンク

魚類と両生類の特徴を併せ持つティクターリクの復元画
進化を否定する創造論者は、分類群間の中間的な特徴を示す化石が得られないことを指して「ミッシング・リンク」と呼んでいる。しかし、分類群間の移行段階と考えられる化石はすでに一部得られている[11][12]。分類群の起源となった種そのものを見つけるのは確かに困難だが、それに近縁な種の化石があれば、進化過程を解明するのに充分である[11]。たとえば爬虫類と鳥類の特徴を併せ持つ化石には有名な始祖鳥に加えて、多数の羽毛恐竜がある[13][14]。クジラの進化過程は、時折水に入る陸生哺乳類であったインドヒウスに始まり、徐々に水中生活に適応していく一連の化石から明らかになっている[15][16]。現在の魚類と両生類をつなぐ移行化石としてはエウステノプテロン、パンデリクチス、アカンソステガ、イクチオステガなどが知られていたが、さらにパンデリクチスよりも両生類に近く、アカンソステガよりも魚類に近いティクターリクが2006年に発表された[17][18]。無脊椎動物では、祖先的なハチの特徴と、より新しく進化したアリの特徴を併せ持つアケボノアリなどの例がある[19]。移行化石は次々と発見されており、たとえば2009年には、鰭脚類(アシカやアザラシ)と陸上食肉類との中間的な特徴を示す化石[20]や、真猿類の祖先に近縁だと考えられるダーウィニウスの化石[21]が報告されている[22]。人類が他の類人猿に似た祖先から進化してくる過程を示す化石も見つかっている[23][24]。
生物地理学から
生物の分布がいかにして成立してきたかを探る分野である生物地理学は、進化を支持する強力な証拠をもたらす。進化生物学者のコインによれば、創造論者は生物地理学上の証拠に反論することができないため、無視を決め込んでいるという[25]。
火山活動などによる海底の隆起によってできた、大陸と繋がったことのない島を海洋島と呼ぶ。ガラパゴス諸島やハワイ、小笠原諸島といった海洋島の在来生物相には海を渡れない両生類、コウモリを除く哺乳類、純淡水魚がほとんど、あるいは全く含まれないのが普通である。それに対して大陸と繋がった歴史のある島には、哺乳類や両生類が普通に分布している。しかも島にすむ生物は、ほとんどの場合最も近い大陸の生物と近縁である。このようなパターンでは、生物が地球の歴史の中でその分布を広げながら進化してきたと考えない限り理解できない[26][27]。
地域が違うと、似たような生息環境であっても異なる生物が分布することがあり、これも進化の証拠となる。同じ砂漠でも新世界にはサボテン科、旧世界にはキョウチクトウ科やトウダイグサ科の乾燥に適応した植物が生息している[28][29]。
ダーウィンの時代には知られていなかったが、地球の歴史上、大陸は長い時間をかけて移動し、離合集散を繰り返してきた(大陸移動説)。生物の分布のなかには、かつて繋がっていた大陸に共通祖先がいて、大陸の分裂に伴って系統が分岐したと考えることでうまく説明できるものも多くある。たとえばシクリッド科の淡水魚や走鳥類の分布は、かつてのゴンドワナ大陸が複数の大陸に分裂した過程で分岐してきたことで成立したと考えられる[30]。
輪状種の存在も、生物がわずかな変化を累積して連続的に進化してきたことの傍証となる。輪状種とは、ある場所では互いに交配せず、別種として区別できる生物が、実は多数の中間型によって連続している場合を指す[31]。ヨーロッパ北西部ではセグロカモメとニシセグロカモメが互いに交配せず別種であると識別できるが、そこから東に向かい、北極の周りを一周してヨーロッパに戻ると、ニシセグロカモメが次第に変化してセグロカモメにいたる一連の亜種が観察でき、明瞭な種の区別はない。
比較解剖学から
相似と相同
進化の証拠は化石だけではなく、現生生物の形態を比較することからも得られている。たとえば陸上脊椎動物は外見上非常に多様であり、コウモリや鳥のように飛翔するものまで含まれる。それにもかかわらず、すべて基本的には同一の骨格を持ち、配置を比較することで相同(進化的な由来を同じくする)な骨を特定することができる。このことは、陸上脊椎動物が単一の共通祖先を持ち、祖先の形態を変化させながら多様化してきたことを示している[32][33]。それぞれの種が独立に誕生したとしたら、鳥の翼と哺乳類の前脚のように全く機能の異なるものを、基本的に同一の骨格の変形のみで作る必然性はない。
機能が異なっていても由来と基本的構造を同じくする相同とは逆に、由来や構造の異なる器官が同一の機能を果たし、類似した形態を持つことを相似という。たとえばコウモリと鳥、翼竜はどれも前肢が翼となっているが、翼を支持する骨は大きく異なっている[34]。鳥は羽毛によって翼の面積を大きくしており、掌や指の骨の多くは癒合して数を減らしているのに対し、コウモリは掌と指の骨を非常に長く発達させて、その間に膜を張ることで翼を構成している。その一方で、翼竜の翼は極端に長く伸びた薬指1本で支持されている。これは、翼を持たなかった共通祖先から、翼を持つ系統がそれぞれ別個に進化してきた(収斂進化)と考えれば合理的に理解できる。
痕跡

ガラパゴスコバネウは飛べないが、痕跡的な翼を持つ。
進化がもともとの形態を改変して進んできたのだとしたら、生物には祖先の形態の名残が見られるはずである。実際に痕跡の例は枚挙に暇がなく、飛べない鳥の持つ痕跡的な翼、洞窟に住むホラアナサンショウウオの痕跡的な眼、ヒトの虫垂などが挙げられる[35][36]。このような現象は、退化と言われ、進化の一側面をなすと考えられる。これらの器官は必ずしも何の機能も持たないわけではないが、本来の機能を果たしていた祖先からの進化を考えない限り、その存在を説明することはできない[35]。
同様の証拠は解剖学のみならず、遺伝子の研究からも得られている。分子生物学の研究により、生物のゲノムには多数の偽遺伝子が含まれることが明らかになった。偽遺伝子とは、機能を持つ遺伝子と配列が似ているにもかかわらず、その機能を失っている塩基配列のことである[3]。偽遺伝子は、かつて機能していた遺伝子が、環境の変化などによって不要になり、機能を失わせる突然変異が自然選択によって排除されなくなったことで生じると考えられている。一例として、嗅覚受容体の遺伝子が挙げられる。多くの哺乳類は嗅覚に強く依存した生活をしているため、多数の嗅覚受容体遺伝子を持つ。しかし視覚への依存が強く嗅覚の重要性が低い霊長類や、水中生活によって嗅覚が必要なくなったイルカ類では、嗅覚受容体遺伝子の多くが偽遺伝子として存在している。これは、霊長類やイルカ類が、より嗅覚に依存する生活をしていた祖先から進化したことを強く示唆している[37]。
不合理な形態
進化は既存の形態を徐々に変化させて進んでいくものであり、一から設計しなおすようなことは起こらない[38]。その結果として機能的に不合理な形態に進化してしまうことがある。極端な例は反回神経である。これは喉頭と脳をつなぐ神経であり、サメではその間を最短に近い経路で結んでいる。しかし、脊椎動物の進化過程で胸や顎の構造が変化するなかで、哺乳類では、この神経は喉頭から心臓の辺りまで下り、その後また上昇して脳にいたるという明らかな遠回りをするようになった。その結果、直線で結べば数センチメートルでよいはずの神経が、ヒトでは10センチメートル程度、キリンでは数メートルに及ぶ長さになっている[39]。同様に哺乳類の輸精管は、精巣とペニスを最短距離で結ぶのではなく、尿管の上まで迂回するように伸びている。これは、哺乳類の進化過程で体内にあった精巣が下に下りてきたときに生じた不合理であると考えられる[40][41]。同様の不合理な形態は、人体にも数多く見られる[42][43]。
系統分類学から
生物分類学の祖とされるリンネはダーウィンより古い時代に生きた創造論者だったが、入れ子状の階層的な分類体系を構築した。生物が共通祖先から分岐を繰り返して多様化してきたものだと考えれば、入れ子の各階層は一つの分岐点を反映するものとして解釈できる。そのため、形態に加えてDNAの塩基配列を含むさまざまな特徴が、例外はあるもののかなり一致した入れ子状の分類体系を支持するという事実は、共通祖先からの進化によって説明できる[44][45]。
近年ではDNAの比較に基づく系統推定が盛んに行われている。このとき、複数の遺伝子をそれぞれ解析すると、細部は異なるにせよおおまかに一致した系統樹を支持することが多い。もし生物がそれぞれ別個に起源していたとしたら、異なる遺伝子が同じ傾向を示すと考える理由はないだろう[46]。
発生生物学から
多細胞生物は一細胞の卵から胚発生の過程を経て体を形成していく。この過程にも、進化の証拠が多く見られる。
有名なのは、ドイツの生物学者エルンスト・ヘッケルの唱えた反復説である。彼は、「個体発生は系統発生を繰り返す」と言われるように、生物は胚発生の過程でその祖先の形態を繰り返すと主張した。現在では、この説は必ずしも成り立たないものとされているが、それでも発生過程に進化の痕跡を見て取れるのは確かである[47][48]。たとえば脊椎動物の胚はすべて魚のような形態をしており、哺乳類のように成体では鰓を持たないものの胚も鰓弓を持つ[47]。
観察された進化

ガラパゴスフィンチの進化は長期の野外調査により観察されている。
家畜化・人為選択も参照。
以上の証拠は過去の進化過程を明らかにするものだが、現在進んでいる進化が観察されたこともある[49]。古典的な例はオオシモフリエダシャクの工業暗化である。このガには白色型と黒色型がいるが、工業の発展に伴う煤煙で樹木表面が黒く汚れた結果、捕食者である鳥から姿を隠しやすい黒色型のガが急激に頻度を増した[50]。次いで有名なのはガラパゴスフィンチの事例で、グラント夫妻らの30年以上にわたる長期の調査により、環境変動に伴う自然淘汰が嘴の進化を引き起こしたことが確認されている[51][52]。病原菌や害虫に抗生物質や殺虫剤で対処しようとすると、急速に薬剤抵抗性が進化することもよく知られている[53]。
実験進化
ロシアの神経細胞学者、リュドミラ・ニコラエブナ・トルットとロシア科学アカデミーの遺伝学者、ドミトリ・ベリャーエフは共同研究でキツネの人為選択による馴致化実験を行った[54][55]。100頭あまりのキツネを掛け合わせ、もっとも人間になつく個体を選択して配合を繰り返すことで、わずか40世代でイヌのようにしっぽを振り、人間になつく個体を生み出すことに成功した。同時に、耳が丸くなるなど飼い犬のような形質を発現することも観察された[56][57][58]。これはなつきやすさという性質が、(自然、あるいは人為的に)選択されうることを示している。
人為的に進化を引き起こす研究も行われている。エンドラーはグッピーを異なる環境に移動させることによって、雄の体色が捕食者と雌による配偶者選択に応じて進化することを明らかにした[59][60]。レンスキーらは大腸菌の長期培養実験によって、代謝能力の進化を観察している[61][62]。また人為淘汰による進化は、農業における品種改良に応用されている[63]。植物では、倍数化による種分化(後述)を実験的に再現することにも成功している[64]。
進化のしくみ
現在、進化を説明する理論として最も支持されているのは進化の総合説と呼ばれるもので、ダーウィンとウォーレスの自然選択説と、メンデルの遺伝子の理論、集団遺伝学の理論や木村資生の中立進化説を統合したものである。この総合説によれば、突然変異によって生じた遺伝子の変異はランダムでない自然選択と、確率的に起こる遺伝的浮動によって個体群中に固定し、新しい形質の出現や種分化などの進化現象を引き起こすと考えられる。
遺伝的変異
ある形質について変異が全くなければ、その形質は進化しない。変異があっても、その変異が次世代に伝わる傾向がなければ(すなわち、遺伝しなければ)進化は起こらない。
DNAの配列に突然変異が生じることで、新たな形質が出現する
遺伝において親から子に受け渡されるのは遺伝子であり、その実体はDNAの塩基配列情報である。DNAは細胞分裂に際して複製されるが、その過程でエラー、すなわち突然変異が起こることがある。これによって生じる個体差が遺伝的変異である。さらには、突然変異によって生じた遺伝子が有性生殖や接合によって組み換えられることによっても、新しい遺伝的変異が生じる[65]。
DNA配列上には現れないが遺伝子発現の変化による遺伝形質の変化についても、研究が進められている。塩基配列の変化を伴わない遺伝子の制御はエピジェネティクスと呼ばれ、DNAのメチル化による遺伝子発現抑制やヒストンの化学修飾による遺伝子発現変化などがある[66]。しかしこの様な化学修飾は細胞分化に大きな役割を持ち、化学修飾が多世代を超えて長期間維持されることはないため、進化の原動力になるか疑問である。ただゲノムには狭義の遺伝子(コーディング領域)のみでなく遺伝子の制御領域(プロモーターやシスエレメント)があり、遺伝子の制御領域の突然変異が進化の原動力になる事がある[67]。
一般的に、突然変異は「ランダム」に起こると言われる。これは、環境に応じて適応的な変異がより生じやすくなるというようなことはない(寒いからといって、毛皮を厚くする突然変異が暑い場所よりも生じやすくなることはないなど)という意味であり、あらゆる意味でランダムというわけではないということに注意が必要である[68][69][70]。ラマルクは、より多く使われた器官が発達し、その発達が次世代に遺伝することで適応的な遺伝的変異が生じるとした(用不用説)が、この説は誤りであることがわかっている[71]。突然変異はこのような説を否定する意味においてのみ「ランダム」である。実際には突然変異はあらゆる意味で「ランダム」とは言えず、たとえば放射線や発癌性物質によって誘発される。
突然変異は発生の過程を変化させることによって表現型を変化させるので、変化の範囲には限りがある[72]。この制約がどの程度実際の進化に影響するかについては議論がある[73]。
このほか、他の生物が持つ遺伝子が他生物に取り込まれることでその遺伝子を獲得することがある。(遺伝子の水平伝播)
遺伝子の頻度変化
遺伝的変異が生じても、その変異(あるいはその変異のもととなる対立遺伝子)を持つ個体が子孫を残さなければ、その変異は個体群から消失する。しかし一部の変異は頻度を増して個体群内に定着(固定)し、個体群の特徴を変化させることになる。
対立遺伝子頻度は、以下の2つの過程によって変化する[74]。
- 自然選択
- 遺伝的浮動
自然選択

自然選択の模式図。図中では色の濃い個体ほど有利とされている。突然変異がさまざまな形質をもたらすが、そのうち生存に好ましくない変異が消滅し、残った個体が次世代に子孫を残す。この繰り返しによって、個体群が進化していく。
一部の遺伝的変異はそれを持つ生物個体の適応度(生存と繁殖)に影響する。その多くは適応度を低下させるので、それを持つ個体は子孫を残せず、変異は消失する(負の自然選択)。しかし、なかには適応度を高める突然変異もある。たとえばレンスキーらは大腸菌の長期培養実験のなかで、クエン酸塩を利用できるようになる突然変異がまれに生じるのを観察した[62]。
適応度を高める対立遺伝子は、それを持つ個体が持たない個体よりも平均して多くの子孫を残すので、個体群内で頻度を増す。この過程を正の自然選択という。正の自然選択によって、生物個体群は世代を経るにつれてより適応的な形質を持つように進化していく。自然選択は、適応進化を説明できる唯一の機構である[75]。

モリマイマイの殻の色彩には大きな変異がある。
自然選択において有利になる形質は環境条件によって異なる。ヨーロッパに生息するカタツムリの一種モリマイマイの殻の色彩は変異が大きく、個体群によって色と模様が異なる。これは、生息環境によって捕食者の目を逃れるのに適した色、体温調節に適した色が異なるため、自然選択によって個体群ごとに異なる色彩が進化したのだと考えられる[76]。形質の適応度がその頻度によって決まることもある。たとえば、もし捕食者が多数派の模様を学習し、まれなタイプの模様はあまり食べないということがあれば、ある模様の適応度がその頻度が少ないときに高くなる。このような自然選択を頻度依存選択と呼ぶ[76]。
広義には自然選択に含まれるが、性選択も適応度に影響する。性選択は、配偶者をめぐる同性間の競争や、異性による配偶者の選り好みによって起こる選択のことをいう。たとえばコクホウジャクという鳥では、長い尾羽を持つ雄が雌に好まれるので、そのような雄の適応度は高くなる[77]。
自然選択は個体あるいは遺伝子を単位として考えられることが多いが、かつては個体の集まったグループを単位とした自然選択(群選択あるいは集団選択)が重視されていた。かつてのような粗雑な群選択理論は今では否定されているが、グループを含む複数の階層での選択を考慮する複数レベル選択説が提唱されており、その重要性について議論になっている[78]。

遺伝的浮動による遺伝子頻度変化のシミュレーション。個体数の少ない集団(上、10個体)では、個体数の多い場合(下、100個体)よりも浮動による変化が大きい。
遺伝的浮動
遺伝的変異のなかには、適応度に全く、あるいはごくわずかしか影響しないものも多い。その場合には、遺伝子頻度はランダムに、確率的に変動することになる。また適応度に影響する場合でも、確率的な変動の影響は受ける。このランダムな遺伝子頻度の変化を遺伝的浮動という[79]。遺伝的浮動はとくに数の少ない個体群において重要である。そのため、少数の個体が新しい生息地に移住して定着した場合に遺伝子頻度が大きく変化することがあり、これを創始者効果という[80]。
木村資生は、遺伝子レベルの進化においては遺伝的浮動が重要であると指摘した(分子進化の中立説)[81]。分子進化の中立説は、塩基配列のデータをよく説明できる。表現型レベルでも、適応度上中立な変化であれば遺伝的浮動によって進化することはありうるが、実際にはほとんどないと考えられている[82](ただし、表現型と分子のそれぞれにおいて、浮動と選択がどの程度重要かについては議論がある[83])。
進化の速度
形態の進化
化石が多く見つかっている系統の進化速度は、より新しい化石と古い化石の形態を比較することで調べることができる。量的な形態進化の速度は、100万年あたりネイピア数倍(約2.7倍)の変化を1ダーウィンとして定義する[84]。離散的な形態の進化については、いくつかの形質状態を定義して、その変化の回数を数えることで計測できる[85]。分類群の数を利用した進化速度の定義もあり、ある期間におけるある系統がいくつの種(あるいは属などより高次の分類群)に分けられるかによって進化速度を測定する。たとえば、ウマ類の系統は現生のものを除くと、5000万年の間に8属を経過してきたため、約625万年あたり1属の進化速度で進化してきたと計算できる[86]。
進化速度は系統によって大きく異なり、進化速度が非常に遅いために祖先の化石種とほとんど変わらない形態を持つものを生きている化石と呼ぶ。ただし、同じ系統でも進化速度は一定ではない。たとえばハイギョ類は生きている化石として有名であり、確かに中生代以降の進化速度はかなり遅いのだが、古生代においてはむしろ急速に進化していた[85]。また、すべての形質の進化速度が同じ傾向を示すわけでもない。ヒトの系統が脳の大きさに関して他の霊長類、たとえばアイアイに比べて急速な進化を遂げてきたのは明らかだが、同時にアイアイの歯はヒトの歯よりも初期霊長類と比べて違いが大きく、歯の形態に関してはアイアイのほうが進化速度が速かったと考えられる[87]。
形態の進化速度に関わる断続平衡説については、種分化との関連で後ほど取り上げる。
分子進化
分子レベルの進化速度は、単位時間(あるいは世代数)あたりの塩基置換数として計測できる。分子進化の中立説によれば、世代あたりの塩基置換速度は中立な突然変異率によって決まるため、突然変異率が一定ならば一定の速度で進化すると予測される。この予測は、塩基配列の比較から系統が分岐した年代を推定する分子時計の根拠となっている[82][88]。
わずかな塩基配列の変化で機能が損なわれるような遺伝子は、中立な突然変異が少ないので、進化速度が遅くなる[89][90]。逆に、もはやその役目を果たさない偽遺伝子ではほとんどの突然変異が中立になるので、進化速度が非常に速い。たとえば、地中に生息し眼が退化したシリアヒメメクラネズミでは、レンズを作るタンパク質をコードする遺伝子が偽遺伝子化し、急速に進化している[91]。
大進化
種内で起こる形質の変化を小進化というのに対し、新しい種や、種より高次の分類群の起源や絶滅のプロセスを大進化という。このような区別がなされるのは、大進化を小進化の積み重ねで説明できるかどうかについて議論があるためである[92]。しかし一般的には、大進化も小進化の延長として理解できると考えられている[92]。
種分化
1種が2種以上に分岐し、新しい種が形成されることを種分化という。種の定義は多数あるが、進化生物学においては「相互に交配可能な生物の集団」として定義されることが多い(生物学的種概念)。したがって種分化は、集団間に生殖隔離が生じることを意味する[93]。
前述したセグロカモメの事例のほか、エシュショルツサンショウウオ[94]などで知られる輪状種の存在は、わずかな進化の累積が種分化を引き起こすことを示している[95]。
一度に種分化が起こる事例も報告されている。たとえばカタツムリの殻の巻きは単一の遺伝子によって決定されているが、この遺伝子に突然変異が起こって右巻きになると、巻きの違う個体同士は交尾できないことが多いので、生殖隔離が成立する[96][97]。植物では、倍数体(全ゲノムが倍化した個体)が、もとの種と生殖できなくなることによる種分化がかなり頻繁に起こっていると考えられている[98]。
断続平衡説

断続平衡説(下)と漸進的進化(上)の対比
地層中の化石の出現パターンを調べると、基本的な形態はあまり変化しないで安定な状態にあり、新しい形態をもつ化石は、ある地層に突然現れ、その後長い年月の間、形態はふたたび安定して、あまり変化しないという傾向がある(ただし、古生物学でいう「突然」とは数万年程度の時間を指す)。古生物学者のエルドリッジとグールドは、このような現象を断続平衡現象と呼んだ[99]。彼らは、進化は種分化のときにのみ急激に起こり、その他の期間は停滞すると主張した。
断続平衡説は種分化の重要な側面を捉えているという評価もある一方で[100]、批判も多い。たとえば断続平衡説は生物学的種概念に基づく種分化の理論を援用しているが、化石種は交配可能性ではなく形態に基づいて分類されているため、化石種と生物学的種は必ずしも一致しない[101]。実際に形態の変化を定量的に追跡できる事例についてみると、断続平衡的な進化を示す系統もあるが、一方で連続的に進化している系統もある[102]。また、断続平衡説は主流の進化理論に矛盾すると言われたこともあるが、実際には一般的な進化理論の範疇で理解できるものである[103][104][105]。
進化に関する誤解
進化という概念は、日常生活でも頻繁に使用されるためか、誤解されていることが多い。よく見られる誤解について以下に述べる。
- 進化に目的があるという誤解
- 「高い所の葉を食べるために、キリンの首は伸びた」といった表現はよく使われる。しかし、首が伸びるという突然変異は目的とは関係なく起こり、それに自然選択が働いたにすぎない[106]。もっとも、無目的な突然変異と自然選択によって、あたかも目的を持って作られたかのような器官が生み出されるのも確かである。ドーキンスはこのことを「盲目の時計職人」という比喩で表現した[107]。
- 進化は進歩であるという誤解
- 地中で生活するモグラの目が退化していることも進化の結果であるように、進化は必ずしも器官の発達や複雑化をもたらすわけではなく、また知能を発達させるとも限らない[108]。まして、ヒトが進化の頂点であり、進化はヒトを目指して進むなどと考えるべきではない[109]。ヒトは他の現生生物と同じく、各々の環境に適応し、枝分かれしながら進む進化系統樹の末端の枝の一つにすぎない。ただしこれは進歩を複雑化やヒト化として定義した場合の話であり、定義によっては何らかの「進歩」(適応の向上、進化可能性の増大など)を進化に見出すことは可能だとする考えもある[110][111]。
- この誤解は、進化(英: evolution)という語にその一因があるかもしれない。英語の「evolution」という語はラテン語の「evolvere」(展開する)に由来し、発生学においては前成説の発生過程を意味する語であったほか、日常語としては進歩(progress)と強く結びついて使われる。そのためダーウィンはこの語をほとんど使わず、変化を伴う由来(descent with modification)という表現を好んだ[112]。evolution という語が生物進化の意味で使われるようになったのはスペンサーの影響である。このことは日本語の「進化」とも関連している。明治初期の日本ではダーウィンよりもスペンサーの影響が大きかったため、evolution は万物の進歩を意味するスペンサーの用語として進化と訳されたが、これが現在にも続く、進化は進歩であるという誤解を招いている[113]。
- 「チンパンジーはいずれヒトに進化するのか」「ヒトがチンパンジーから進化したなら、なぜチンパンジーがまだいるのか」という疑問
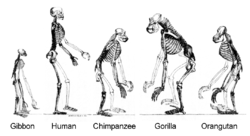
ヒトと他の類人猿は、共通の祖先から進化した
- ヒトはチンパンジーと共通祖先を持ち、ヒトもチンパンジーもそこから独自の進化を遂げてきたにすぎないため、この疑問は的外れである[114][115][116]。ヒトは数百万年前に、チンパンジー(およびボノボ)との共通祖先から分岐したと推定されている。この共通祖先はたまたまヒトよりはチンパンジーに似ていたと思われるが、それはヒトの系統がより多くの変化を遂げた結果にすぎず、共通祖先はチンパンジーともヒトとも異なる類人猿であった[115]。
- この誤解は、生物は下等なものから高等なものまで一列に配列され、進化はその序列の中で梯子を登るように進むという、より深い誤解を反映したものである[114]。実際には、進化は分岐を繰り返しながら進むものであり、現生の生物はどれも等しく系統樹の末端に位置づけられる。
生物学以外での「進化」概念
「進化」という概念は、ダーウィン以来の進化生物学の成功により有力となったが、生物学の影響を受けて、あるいはそれとは独立に「進化」という概念は、さまざまな学問分野において重要な役割を果たしている。たとえば、「進化経済学」[117]「進化経営学」「進化心理学」「進化的計算」などは前者の例、「宇宙の進化」[118]は後者の例である。
生物学の影響を受け、「進化」概念を研究・分析の中核に据えるとき、進化生物学の進化概念をどの程度忠実に移植するかについての議論は多い[117]。進化経済学では、意図せざる進化と共に、意図された進化が重要であるとされることが多い[119]。
脚注
- ^ abcRidley(2004) p.4
- ^ abcFutuyma(2005) p.2
- ^ abcde『岩波生物学辞典』
- ^ abソーバー(2009) pp.7-8
- ^ abコイン(2010)
- ^ abドーキンス(2009)
^ Futuyma(2005) p.2
^ コイン(2010) pp.66-71
^ Ridley(2004) p.64
- ^ abコイン(2010) pp.72-78
- ^ abコイン(2010) pp.78-81
^ ドーキンス(2009) Ch.6
^ コイン(2010) pp.87-98
^ Chiappe(2009)
^ コイン(2010) pp.98-105
^ Thewissen et al.(2009)
^ ドーキンス(2009) pp.230-261
^ シュービン(2008) Ch.2
^ コイン(2010) pp.106-107
^ Rybczynski et al.(2009)
^ Franzen et al.(2009)
^ ドーキンス(2009) p.266, pp.275-276
^ コイン(2010) Ch.8
^ ドーキンス(2009) Ch.7
^ コイン(2010) p.164
^ コイン(2010) pp.181-197
^ Futuyma(2005) p.119
^ コイン(2010) pp.168-169
^ Futuyma(2005) p.118
^ Futuyma(2005) pp.126-128
^ パターソン(2001) pp.8-11
^ 遠藤(2006)
^ ドーキンス(2009) pp.409-422
^ 遠藤(2006) pp.109-117
- ^ abコイン(2010) pp.112-128
^ ドーキンス(2009) pp.481-483, pp.490-493
^ コイン(2010) pp.133-136
^ 遠藤(2006) pp.47-48
^ ドーキンス(2009) pp.501-507
^ ウィリアムズ(1998) pp.232-236
^ ドーキンス(2009) pp.507-508
^ 遠藤(2006) Ch.4
^ ドーキンス(2009) p.508
^ コイン(2010) pp.38-41
^ Ridley(2004) pp.61-63
^ ドーキンス(2009) pp.455-457
- ^ abコイン(2010) pp.139-151
^ 倉谷(2005)
^ Ridley(2004) pp.45-47
^ Majerus(2009)
^ ワイナー(1995)
^ Grant & Grant(2002)
^ ワイナー(1995) Ch.18
^ 動物好きな研究者の夢 -- 40年の研究からペットギツネが誕生
^ 実験飼育場で遊ぶキツネ
^ 地球ドラマチック「不自然な“進化”~今 動物に何が!?~」
^ 特集:野生動物 ペットへの道
^ ロシア科学アカデミーシベリア支部 細胞学・遺伝学研究所の「キツネの家畜化研究」
^ ワイナー(1995) pp.119-126
^ ドーキンス(2009) pp.216-224
^ ドーキンス(2009) pp.194-216
- ^ abBlount et al.(2008)
^ Ridley(2004) p.47
^ Ridley(2004) pp.53-54
^ Ridley(2004) pp.87-88
^ 佐々木(2005)
^ Jiménez-Delgado S et al (2009)
^ ドーキンス(2004a) pp.482-485, 493-494
^ Ridley(2004) p.88-89
^ Futuyma(2005) pp.178-179
^ ドーキンス(2004a) pp.452-453
^ ドーキンス(2004a) pp.491-493
^ ルース(2008) Ch.11
^ コイン(2010) pp.46-47
^ ドーキンス(2004a) p.454
- ^ abパターソン(2001) pp.96-99
^ Andersson(1982)
^ 松本(2010)
^ Futuyma(2005) p.226
^ Futuyma(2005) pp.231-232
^ 木村(1988) pp.54-55
- ^ abFutuyma(2005) p.236
^ 斎藤(2008)は表現型の進化も浮動によって起こる可能性を指摘しているが、逆にオール(2009)は分子進化も相当部分が選択によると主張している。
^ Ridley(2004) p.591
- ^ abRidley(2004) pp.606-607
^ シンプソン(1977) pp.102-103
^ シンプソン(1977) p.104
^ パターソン(2001) p.110-111
^ Futuyma(2005) pp.236-237
^ パターソン(2001) p.117
^ Hendriks et al.(1987)
- ^ ab河田(1990) pp.114-115
^ Ridley(2004) p.381
^ Moritz et al.(2002)
^ Ridley(2004) pp.50-53
^ Ueshima & Asami(2003)
^ Hoso et al.(2010)
^ コイン(2010) pp.321-326
^ 河田(1990) pp.124-125
^ ステルレルニー(2004) pp.98-99
^ 河田(1990) pp.127-128
^ Ridley(2004) pp.605-606
^ ステルレルニー(2004) p.98
^ Ridley(2004) pp.600-601
^ ドーキンス(2004a) p.402
^ 長谷川・長谷川(2000) pp.36-37
^ ドーキンス(2004a) p.46
^ 長谷川・長谷川(2000) pp.39-40
^ 長谷川・長谷川(2000) p.38
^ ステルレルニー(2004) pp.140-143
^ ドーキンス(2004b)
^ グールド(1984)
^ 松永(2009) pp.51-52
- ^ ab長谷川・長谷川(2000) pp.40-43
- ^ abドーキンス(2006) pp.159-160
^ 渡辺(2010) p.68
- ^ ab『進化経済学ハンドブック』「概説」1.「経済における進化」
^ 海部・吉岡(2011)
^ Ziman(2000), I. Evolutionary thinking
参考文献
Andersson, Malte (1982年). “Female choice selects for extreme tail length in a widowbird”. Nature 299 (5886): 818-820. doi:10.1038/299818a0.
Blount, Zachary D.; Borland, Christina Z.; Lenski, Richard E. (2008年). “Historical contingency and the evolution of a key innovation in an experimental population of Escherichia coli”. Proceedings of the National Academy of Sciences of the United States of America 105 (23): 7899-7906. doi:10.1073/pnas.0803151105. PMID 18524956.
Chiappe, Luis M. (2009年). “Downsized dinosaurs:the evolutionary transition to modern birds”. Evolution:Education and Outreach 2 (2): 248-256. doi:10.1007/s12052-009-0133-4.
- コイン, ジェリー・A 『進化のなぜを解明する』 塩原通緒訳、日経BP社、2010年(原著2009年)。ISBN 9784822284206。
- ドーキンス, リチャード 『盲目の時計職人』 日高敏隆監修、中嶋康裕・遠藤彰・遠藤知二・疋田努訳、早川書房、2004年a(原著1986年)。ISBN 4152085576。
- ドーキンス, リチャード 「人間至上主義と進化的な進歩」『悪魔に仕える牧師』 垂水雄二訳、早川書房、2004年b(原著2003年)、363-382頁。ISBN 4152085657。
- ドーキンス, リチャード 『祖先の物語』上、垂水雄二訳、小学館、2006年(原著2004年)。ISBN 4093562113。
- ドーキンス, リチャード 『進化の存在証明』 垂水雄二訳、早川書房、2009年(原著2009年)。ISBN 9784152090904。
- 遠藤秀紀 『人体 失敗の進化史』 光文社〈光文社新書〉、2006年。ISBN 433403358X。
Franzen, Jens L.; Gingerich, Philip D.; Habersetzer, Jörg; Hurum, Jørn H.; von Koenigswald, Wighart; Smith, B. Holly (2009年). “Complete primate skeleton from the middle Eocene of Messel in Germany:morphology and paleobiology”. PLoS ONE 4 (5): e5723. doi:10.1371/journal.pone.0005723. PMID 19492084.
Futuyma, Douglas J. (2005). Evolution. Sinauer Associates. ISBN 0878931872.
- グールド, スティーヴン・ジェイ 「ダーウィンのジレンマ―「エヴォリューション」の長い旅」『ダーウィン以来』上、浦本昌紀・寺田鴻訳、早川書房、1984年(原著1977年)、42-49頁。ISBN 415203243X。
Grant, Peter R.; Grant, B. Rosemary (2002年). “Unpredictable evolution in a 30-year study of Darwin's finches”. Science 296 (5568): 707-711. doi:10.1126/science.1070315.
- 長谷川寿一、長谷川眞理子 『進化と人間行動』 東京大学出版会、2000年。ISBN 4130120328。
Hendriks, W.; Leunissen, J.; Nevo, E.; Bloemendal, H.; de Jong, W. W. (1987年). “The lens protein alpha A-crystallin of the blind mole rat, Spalax ehrenbergi:evolutionary change and functional constraints”. Proceedings of the National Academy of Sciences of the United States of America 84 (15): 5320-5324. PMID 3474658. http://www.pnas.org/content/84/15/5320.short.
- 海部宣男・吉岡 一男 『進化する宇宙』 放送大学教育振興会、2010年、新訂。ISBN 978-4595312885。
Hoso, Masaki; Kameda, Yuichi; Wu, Shu-Ping; Asami, Takahiro; Kato, Makoto; Hori, Michio (2010年). “A speciation gene for left-right reversal in snails results in anti-predator adaptation”. Nature Communications 1: 133. doi:10.1038/ncomms1133. http://www.natureasia.com/ja-jp/ncomms/abstracts/35003.
- 河田雅圭 『はじめての進化論』 講談社〈講談社現代新書〉、1990年。ISBN 4061489836。
- 木村資生 『生物進化を考える』 岩波書店〈岩波新書〉、1988年。ISBN 4004300193。
- 倉谷滋 『個体発生は進化をくりかえすのか』 岩波書店〈岩波科学ライブラリー108〉、2005年。ISBN 4000074482。
- 松本俊吉 「自然選択の単位の問題」『進化論はなぜ哲学の問題になるのか』 松本俊吉編著、勁草書房、2010年、1-25頁。ISBN 9784326101986。
- 松永俊男「日本におけるダーウィン理解の誤り」、『現代思想』第37巻第5号、2009年、 48-52頁、 NAID 40016546740。
Majerus, Michael E. N. (2009年). “Industrial melanism in the peppered moth, Biston betularia:an excellent teaching example of Darwinian evolution in action”. Evolution:Education and Outreach 2 (1): 63-74. doi:10.1007/s12052-008-0107-y.
Moritz, Craig; Schneider, Christopher J.; Wake, David B. (2002年). “Evolutionary relationships within the Ensatina eschscholtzii complex confirm the ring species interpretation”. Systematic Biology 41 (3): 273-291. doi:10.1093/sysbio/41.3.273.
- オール, H.A.「ゲノムから見た自然選択のパワー」、『日経サイエンス』第39巻第4号、2009年、 26-35頁、 ISSN 0917009X、 NAID 40016490702。
- パターソン, コリン 『現代進化学入門』 馬渡峻輔・上原真澄・磯野直秀訳、岩波書店、2001年(原著1999年)。ISBN 4000053175。
Ridley, Mark (2004). Evolution (3rd ed.). Blackwell Publishing. ISBN 1405103450.
- ルース, マイケル 『ダーウィンとデザイン』 佐倉統・土明文・矢島壮平訳、共立出版、2008年(原著2003年)。ISBN 9784320056640。
Rybczynski, Natalia; Dawson, Mary R.; Tedford, Richard H. (2009年). “A semi-aquatic Arctic mammalian carnivore from the Miocene epoch and origin ofPinnipedia”. Nature 458: 1021-1024. doi:10.1038/nature07985.
- 斎藤成也「遺伝子から見た人間進化」、『科学』第78巻第12号、2008年、 1338-1343頁、 ISSN 00227625、 NAID 40016347408。
- 佐々木裕之 『エピジェネティクス入門』 岩波書店〈岩波科学ライブラリー101〉、2005年。ISBN 4000074415。
Jiménez-Delgado, Senda; Pascual-Anaya, Juan; Garcia-Fernàndez, Jordi (2009年). “Implications of duplicated cis-regulatory elements in the evolution of metazoans:the DDI model or how simplicity begets novelty”. Briefings in Functional Genomics and Proteomics 8: 266-75. doi:10.1093/bfgp/elp029. PMID 19651705.
- シュービン, ニール 『ヒトの中の魚、魚の中のヒト』 垂水雄二訳、早川書房、2008年(原著2008年)。ISBN 9784152089557。
- シンプソン, ジョージ・ゲイロード 『進化の意味』 平沢一夫・鈴木邦雄訳、草思社、1977年(原著1967年)。ISBN 4794200560。
- ソーバー, エリオット 『進化論の射程』 丹治信春監修、松本俊吉・網谷裕一・森元良太訳、春秋社〈現代哲学への招待〉、2009年(原著2000年)。ISBN 9784393323182。
- ステルレルニー, キム 『ドーキンスvs.グールド』 狩野秀之訳、筑摩書房〈ちくま学芸文庫〉、2004年(原著2001年)。ISBN 4480088784。
Thewissen, J. G. M.; Cooper, Lisa Noelle; George, John C.; Bajpai, Sunil (2009年). “From land to water:the origin of whales, dolphins, and porpoises” (PDF). Evolution:Education and Outreach 2 (2): 272-286. doi:10.1007/s12052-009-0135-2. http://link.springer.com/content/pdf/10.1007%2Fs12052-009-0135-2.
Ueshima, Rei; Asami, Takahiro (2003年). “Single-gene speciation by left-right reversal”. Nature 425 (6959): 679. doi:10.1038/425679a.
- 渡辺政隆「社会の中の進化論」、『学術の動向』第15巻第3号、2010年、 67-70頁、 doi:10.5363/tits.15.3_67、 ISSN 13423363、 NAID 40017030387。
- ワイナー, ジョナサン 『フィンチの嘴』 樋口広芳・黒沢令子訳、早川書房、1995年(原著1994年)。ISBN 4152079487。
- ウィリアムズ, ジョージ・C 『生物はなぜ進化するのか』 長谷川眞理子訳、草思社〈サイエンス・マスターズ9〉、1998年(原著1997年)。ISBN 479420809X。
- 「進化」「偽遺伝子」「退化」「退行的進化」『岩波生物学辞典』 八杉龍一・小関治男・古谷雅樹・日高敏隆編集、岩波書店、1998年、第4版、CD-ROM版。
Ziman, John (2003). Technological Innovation as an Evolutionary Process. Cambridge Uninversity Press. ISBN 9780521542173.
関連項目
- ティンバーゲンの4つのなぜ
- ミーム
外部リンク
- 〈進化〉って何だろう?|JSTバーチャル科学館
Evolution (英語) - Encyclopedia of Earth「進化」の項目。
Evolution (英語) - インターネット哲学百科事典「進化」の項目。
Evolution (英語) - スタンフォード哲学百科事典「進化」の項目。
| ||||||||||||||||||||
| ||||||||||||||


